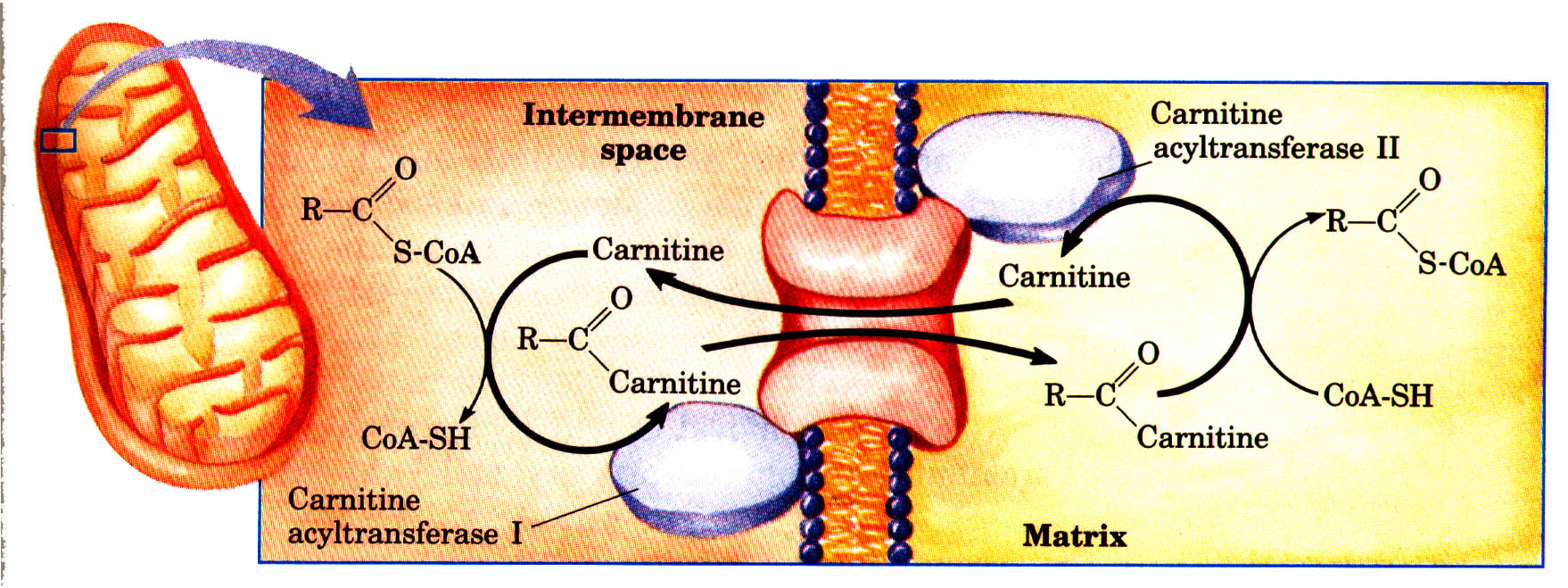
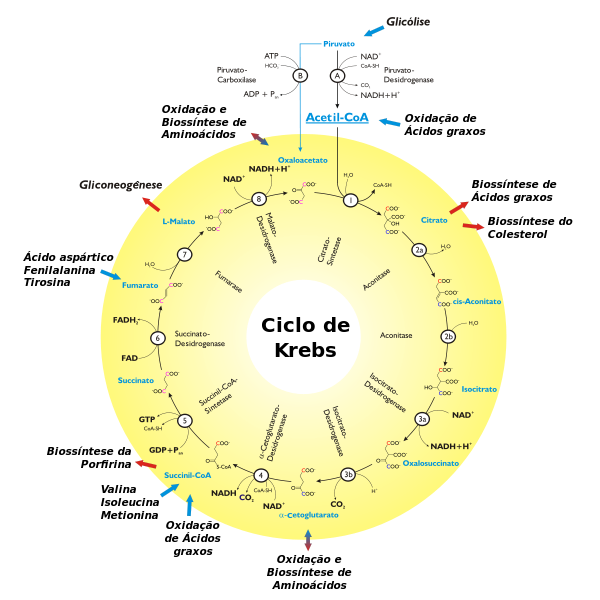
Ainda na segunda semana de aula falamos sobre as proteínas e seu envolvimento no metabolismo, e para entende-las melhor devemos compreender a funcionalidade e o que determina sua síntese. As
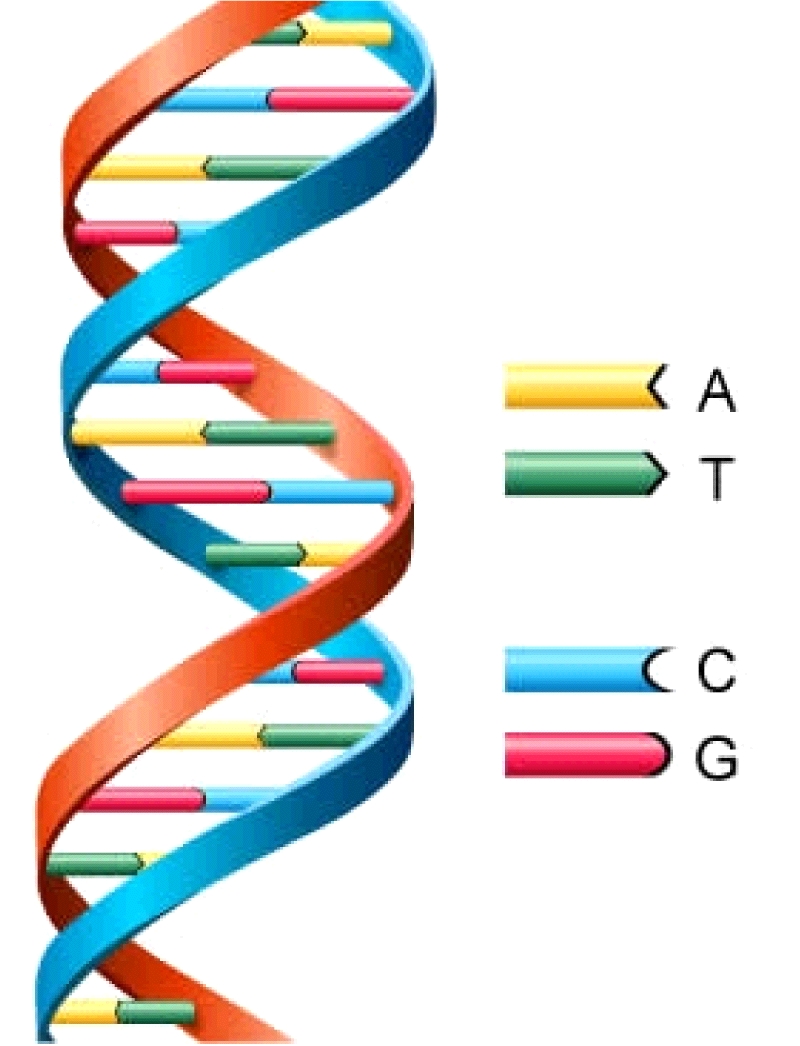
informações genéticas do ser humano estão contidas no DNA, mas qual a relação
com as proteínas? É ele que vai determinar o tipo e a quantidade de proteínas que
serão sintetizadas. O DNA é uma molécula de fita dupla, composto por bases
nitrogenadas (adenina, timina, guanina e citosina). A cada trinca de bases
nitrogenadas, por exemplo, adenina, guanina e timina dão o nome de gene. O
mesmo é responsável pela síntese proteica, porém no DNA só 3% dos genes são
gênicos, os outros 97% não.




Referências:
Maughan, R; Gleeson, M; Greenhaff, P. Bioquímica do Exercício e do Treinamento. São Paulo: Manole, 2000. 240 p. : il.
Diferente dos carboidratos e dos lipídios, as proteínas tem função estrutural, ou seja, não são armazenadas. Além disso desempenham a função de ser base estrutural de todos os tecidos e órgãos. Juntamente com aminoácidos, que falaremos um pouco mais a frente, é precursora de hormônios, peptídios reguladores e neurotransmissores.
Os aminoácidos são responsáveis pela formação das proteínas. Os peptídeos são a junção de aminoácidos, entre 70 e 100 aminoácidos. Já as proteínas são o composto formado por mais de 100 aminoácidos. Todos estes aminoácidos são compostos por carbono, hidrogênio e oxigênio, assim como os outros dois macronutrientes vistos nas outras postagens, no entanto os aminoácidos apresentam como diferencial nitrogênio e dois deles enxofre. E como podemos ver na imagem abaixo os aminoácidos são formados por um grupo amina (NH2) e um grupo carboxila (COOH) ligados a um carbono.
No metabolismo as proteínas exercem funções específicas e
dentre elas ressaltaremos o ciclo alanina-glicose. Os aminoácidos também são
usados como fontes de energia para o metabolismo oxidativo, podendo participar
da síntese da glicose ou de corpos cetônicos. Quando há privação alimentar a
síntese da glicose é produzida por esta via, que torna portanto as proteínas
intramusculares prontas para a degradação. Há portanto síntese
da alanina pelo processo de transaminação que ocorre devido ao piruvato oriundo
da glicose. A partir daí a alanina sai do músculo percorrendo a corrente
sanguínea, até chegar no fígado, onde ocorre a desaminação (perda do grupo
amina) que o transforma em piruvato e, consequentemente, glicose. Assim a
glicose é jogada à corrente sanguínea novamente até chegar ao músculo.

Além disso, em aula vimos um gráfico referente ao suor (nitrogênio e ureia) em relação a diferentes momentos: em repouso, exercício e exercício com baixa reserva de glicogênio. Podemos ver abaixo o gráfico que nos mostra os resultados:

Referências:
Maughan, R; Gleeson, M; Greenhaff, P. Bioquímica do Exercício e do Treinamento. São Paulo: Manole, 2000. 240 p. : il.