Na segunda semana vimos a composição dos lipídios e a β-oxidação. Lipídios (do grego lipos = gordura) são moléculas que possuem os mesmos elementos estruturais que os carboidratos, visto no post anterior, diferenciando-se somente nos tipos de ligações e número de átomos. Podem ser de fonte vegetal ou animal e são divididos em lipídios simples, lipídios compostos e lipídios derivados.


Os lipídios simples, consistem nos triglicerídios e são a principal forma de armazenamento de gordura nos adipócitos. Uma molécula de triglicerídio é formada por um um glicerol ligado a três ácidos graxos. Quando comparada às proteínas e aos carboidratos, a gordura armazenada proporciona quantidades quase ilimitadas de energia. Em um homem adulto jovem típico, as reservas de energia provenientes de gorduras provêm de duas principais fontes: 3.000 kCal dos triglicerídios intramusculares (12 mmol/kg de músculo), que estão em grande proximidade com as mitocôndrias (predominante nas fibras de contração lenta) e entre 60.000 e 100.000 kCal dos triglicerídios dos adipócitos, que mobilizarão os ácidos graxos. Antes da energia ser liberada pelas gordura, a molécula de triglicerídio sofre lipólise no citoplasma, sob a ação da HSL resultando em uma molécula de glicerol e três moléculas de ácidos graxos, como na reação abaixo:
Triglicerídio + 3 H2O -----------> Glicerol + 3 Ácidos graxos
Após a HSL estimular a difusão dos ácidos graxos do adipócito para a circulação, eles irão, em sua maioria, combinar com albumina plasmática para serem transportados para os tecidos ativos sob a forma de AGL (ácidos graxos livres). Na área muscular, em específico no sarcoplamsa, os AGL são liberados para serem transportados por difusão facilitada por um sistema carreador através da membrana plasmática. Dentro da fibra muscular os AGL desempenham as funções de serem reesterificados em triglicerídios, e combinar-se com proteínas para penetrarem nas mitocôndrias participando do metabolismo energético pela ação da carnitina aciltransferase. A carnitina aciltranferase, localizada na superfície externa da membrana mitocondrial (CAT I), catalisa a transferência de um grupo acil (palmitato) para a carnitina, formando acilcarnitina (neste caso usando o exemplo da molécula de acil palmitato). Por sua vez, a superfície interna da membrana mitocondrial (CAT II) regenera acil-CoA e carnitina livre.

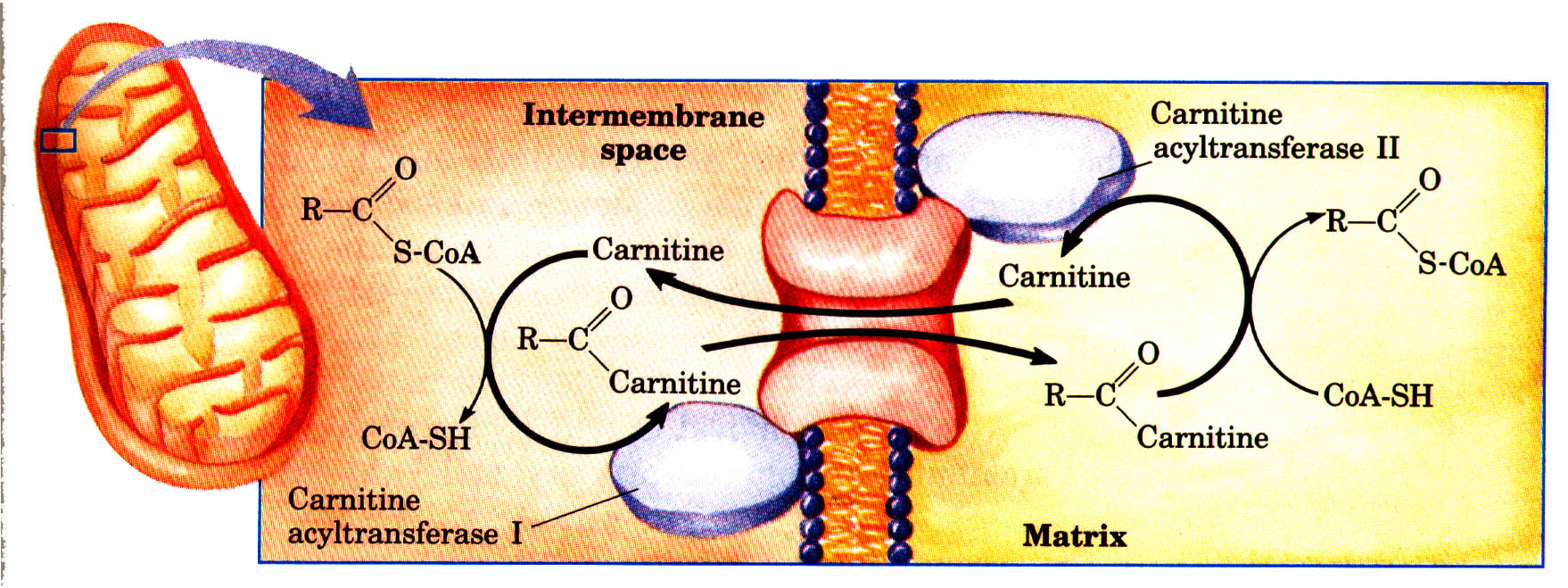
Essa molécula de acil-palmitato existente agora dentro da mitocôndria dará origem a β-oxidação. Consistente na remoção de duas unidades de 2-C (dois carbonos) da cadeia do ácido graxo (acil-palmitato, 16 carbonos), na forma de AcCoA, dando início ao Ciclo de Krebs. Logo, podemos identificar que a cada quebra da ligação beta (segunda ligação entre os carbonos) há a liberação de AcCoA, NADH, FADH. Cada quebra vai liberar 5 ATP, e uma AcCoA. Os resultados são: gasto de dois ATP para a ativação do processo, produto de 35 ATP (devido as sete quebras) oriundo da β- oxidação e o produto de 96 ATP (devido a produção de 8 AcCoA que gera, consequentemente 8 ciclos de Krebs, cada um gerando 12 ATP) oriundo do ciclo de krebs, totalizando 129 moléculas de ATP a cada oxidação completa de uma molécula de palmitato.


A produção de AcCoA formada durante a β-oxidação irá entrar no Ciclo de Krebs ou cadeia respiratória, desde que haja oxaloacetato para a produção de citrato. No entanto quando não há grande disponibilidade de oxaloacetato ou uso do sistema lipídico se sobressaindo com relação ao glicolítico, a AcCoA é enviada ao fígado (único órgão capaz de sintetizar cetonas) para formar cetonas. Isso se dá em momentos de exercício físico duradouro, jejum, diabetes mellitus. Essa produção de corpos cetônicos é decorrente da união de AcCoA+AcCoA. As cetonas são utilizadas quando captadas pelo miocárdio, rins e cérebro sendo usadas como fonte de energia alternativa quando apresentamos a baixa disponibilidade do macronutriente carboidrato ou para auxiliar na concentração de glicose sanguínea.

Além disso, vimos que uma dieta rica em carboidratos auxilia no processo de retardar a instalação da fatiga. Assim para podermos ter o sistema lipolítico funcionando corretamente é bom termos o sistema glicolítico abastecido, pois podemos fornecer mais piruvato e assim formar oxalacetato e todo o ciclo de krebs. Em exercícios prolongados os lipídios poderão fornecer em torno de 80% da energia necessária, tendo como explicação a queda da concentração de glicose no sangue e aumento da liberação de glucagon.

O gráfico visto em aula nos mostra exatamente isso: uma dieta rica em carboidratos é muito mais eficaz em caso de exercícios de longa duração, pois como podemos ver, quanto maior a reserva de glicogênio no sangue maior será o tempo até a instalação da fadiga.
Referências:
William D. McArdle, Frank I. Katch, Victor L Katch, traduzido por Giuseppe Taranto. Fisiologia do exercício: nutrição, energia e desempenho humano. Rio de Janeiro: Guanabara Koogan, 2011.
Maughan, R; Gleeson, M; Greenhaff, P. Bioquímica do Exercício e do Treinamento. São Paulo: Manole, 2000. 240 p. : il.
Nenhum comentário:
Postar um comentário